Глава 2. Значение и формирование рас
Формирование рас, разновидностей и пород
Разновидности в животном мире
Таксономии рас
Расовые различия в заболеваемости
Существуют ли расы?
Имея дело с расовыми различиями в интеллекте необходимо определить как интеллект, так и расу. В предыдущей главе был определён интеллект, в этой главе предлагается определение расы. Простое и прямое определение состоит в том, что раса является группой, которая заметно отличается от других групп. Более полное определение: раса — это воспроизводящаяся популяция, которая в определённой степени генетически отлична от соседних популяций, что возникает в результате географической изоляции, культурных факторов и эндогамии, демонстрирующая характерные наборы генотипических различий в частотах ряда интеркоррелированных, генетически детерминированных характеристик при сравнении с другими воспроизводящимися популяциями. В зонах географического контакта между расами обычно имеются межрасовые гибриды, для которых характерны промежуточные значения частот встречаемости гена в сравнении центральными распределениями в смешивающихся группах. Эти гибриды и популяции смешанных рас известны как переходные формы или клины.
Формирование рас, разновидностей и пород
Общим принципом эволюционной биологии является то, что когда популяции биологического вида оказываются изолированными друг от друга, они эволюционируют в два или более подвида. Эти подвиды обычно называют разновидностями, штаммами или породами. В случае человека разновидности называют расами. Эти разновидности развиваются в результате четырех процессов, а именно эффекта основателя, дрейфа генов, мутаций и адаптации. Эффект основателя заключается в том, что когда популяция разделяется и одна из групп мигрирует к новому местообитанию, то группа, которая мигрирует, не будет генетически идентична оставшейся на прежнем месте. Следовательно, эти две популяции будут отличаются генетически. Эффект дрейфа генов состоит в том, что частоты встречаемости генов изменяются с течением времени до некоторой степени в случайном порядке, и это приводит к различиям между популяциями. Дрейф продолжается, и со временем приводит к увеличению различий между расами. Влияние мутаций заключается в том, что новые аллели (аллели — альтернативные формы генов) появляются в популяциях в случайном порядке, и если они благоприятны для выживания и воспроизводства, то будут постепенно распространяться в популяции. Благоприятная новая аллель может появиться в результате мутации в одной расе, а в других нет. Эффект адаптации состоит в том, что после миграции популяции к новому местообитанию некоторые аллели, не являвшиеся благоприятными на старом местообитании, становятся благоприятными. Особи, обладающие благоприятными аллелями, на новом местообитании производят больше жизнеспособного потомства, так что их аллели будут отбираться и постепенно распространяться в популяции. Новые разновидности нескольких видов развились как адаптивные, когда их популяции мигрировали в арктическую зону. У некоторых из них, таких как лисы, медведи или зайцы, образовался белый мех, который маскирует их, и они не так легко могут быть замечены хищниками или добычей. Во всех этих случаях белый мех появился вследствие мутаций и распространился в популяции потому, что давал животным преимущество в процессе отбора. В конечном итоге, новые благоприятные аллели полностью заменяют менее благоприятные и, как тогда говорят, они становятся «закреплёнными».
Во многих случаях неясно, почему разные разы эволюционировали по тем или иным признакам. Например, мех у европейской белки рыжий, в то время как североамериканские белки серые. Возможно, один из этих цветов даёт преимущество в процессе отбора, и появился случайно в одной из популяций в результате генетической мутации.
Разновидности в животном мире
Было давно установлено, что большинство биологических видов имеют несколько разновидностей, которые в случае человека называют расами. В начале своей карьеры Чарльз Дарвин обратил внимание на различные варианты черепах на Галапагосских островах, и именно это заставило его задуматься о том, как они развились. Позже, в книге «Изменение домашних животных и культурных растений»(Darwin, 1868), он описал породы многих биологических видов, в частности породы голубей, каждая из которых имеет свою особенную манеру полёта, движения и воркования.
Существует много разновидностей или рас среди обезьян. Имеется четыре расы шимпанзе. Это истинный шимпанзе (Pan satyrus verus), обитающий в западной Африке между Гвинеей и Нигерией, лысый шимпанзе (Pan satyrus satyrus) из Камеруна и Габона, карликовый шимпанзе (Pan satyrus paniscus) с севера центрального Заира, и шимпанзе Швейнфурта (Pan satyrus schweinfurthi) из северо-восточного Заира. Эти расы отличаются по физическому облику, по распределению групп крови и по крикам, которые они издают. Различные расы развились среди животных видов в соответствии с теми же принципами, что и у человека. Например, существует две расы горилл. Это горная горилла (Gorilla beringei), обитающая в горах вокруг озёр Эдвард и Киву в восточном Заире, Руанде и западной Уганде, и береговая горилла (Gorilla gorilla)из лесов Камеруна и Габона. Эти две расы географически отделены друг от друга расстоянием примерно в тысячу миль и у них развились различия в физическом облике и группе крови. У горной гориллы более узкий череп, более короткие руки, более длинные ноги, более толстые волосы и группа крови А, в то время как у береговой гориллы череп шире, руки длиннее, ноги короче, волосы тоньше и группа крови В (Baker, 1974). Некоторые из различий между двумя расами развились в ходе адаптации к особенностям среды обитания. Горная горилла населяет более холодную и открытую местность, в то время как береговая горилла населяет более теплую и густо покрытую лесом территорию. У горных горилл развились более толстые волосы, чем у горилл побережья, для защиты от холода. У береговых горилл развились более длинные руки, чтобы перепрыгивать с дерева на дерево. Нет очевидного объяснения тому, почему у горной гориллы более узкий череп, более длинные ноги и группа крови А. Эти различия возникли, возможно, вследствие эффекта основателя, дрейфа генов или случайных мутаций, или же они могут давать животным какие-то неизвестные нам преимущества.
Существует также множество разновидностей домашних животных. Они обычно называются породами и были выведены человеком в различных практических целях. Часто породы выводились с целью увеличения размера животных или, в случае рогатого скота, надоев молока. В некоторых случаях выводились породы, лучше приспособленные к определённой среде обитания. Например, для выпаса на горных пастбищах были выведены породы выносливых овец, отличающиеся от овец низменных местностей. Человеком выведено целых семьдесят девять пород собак с различными способностями, такие как ретриверы для игр, овчарки как помощники в выпасе овец, ротвейлеры для охраны жилища, кокер-спаниели как домашние любимцы и так далее. Эти породы отличаются по общему интеллекту, по специфическим способностям и по лёгкости дрессировки (Согеп, 1994).
Таксономии рас
Биологи и антропологи начали анализировать и классифицировать расы в середине восемнадцатого столетия. Первая таксономия рас была выработана шведским биологом Карлом Линнеем в 1758 г. В своей «Системе природы» он описал четыре человеческие расы, которые он назвал Europaeus(европейцы), Afer(чёрные африканцы), Asiaticus(азиаты) и Americanus(американские индейцы). В 1776 г. немецкий врач Иоганн Фридрих Блюменбах добавил пятую расу и предложил классификацию, основанную преимущественно на цвете кожи. Он называл эти расы кавказская (белая), монгольская (жёлтая), эфиопская (чёрная), американская (красная) и малайская (коричневая). Эти таксономии были основаны на объединении морфологических особенностей и цвета кожи разных рас, как то белая кожа, прямые волосы и узкий нос у европейцев, чёрная кожа, вьющиеся волосы и широкий нос у африканцев, тёмные волосы, желтоватая кожа и уплощённый нос у монголов (аборигенов Восточной Азии), красноватая кожа и клювообразный нос у американских индейцев, и коричневая кожа у малазийцев. Мортон (Morton, 1849) использовал пяти-расовую классификацию Блюменбаха, когда он выполнил первый сравнительный анализ размера мозга у разных рас.
В начале двадцатого столетия были собраны данные о различиях в частотах групп крови человека в разных популяциях всего мира. Хиршфельд и Хиршфельд (Hirszfeld, Hirszfeld, 1919) показали, что частоты некоторых групп крови связаны с расовыми различиями в окраске кожи и в морфологии. Например, группа крови А присутствует у 41–48 процентов европейцев, но только приблизительно у 28 процентов африканцев района Африки южнее Сахары, в то время как группа крови В наличествует у 10–20 процентов европейцев и приблизительно у 34 процентов африканцев. У американских индейцев практически не встречаются группы крови А или В, у них почти у всех группа крови 0.
Данные о распределении групп крови по резус-фактору (Rh) были использованы Бойдом (Boyd, 1950) для следующей пяти-расовой таксономии: (1) Европейцы с высокими частотами групп крови Rh cde и cde; (2) Африканцы с высокими частотами Rh cde; (3) Аборигены Восточной Азии с высокой частотой В и фактическим отсутствием cde; (4) Американские индейцы с очень высокой частотой 0, отсутствием В и редкой cde и (5) Австралоиды с высокой частотой А и крайне редкими В и cde. Этот анализ показал, что распределение групп крови согласуется с расовыми таксономиями классической антропологии по морфологии и окраске кожи.
Более детальная таксономия рас была разработана Куном, Гарном и Бёрдселлом (Coon, Gam & Birdsell, 1950), они предложили рассматривать семь основных рас, каждая из которых подразделяется на две или более подрас. По их классификации основными расами являются: (1) кавказоиды, подразделяющиеся на нордиков Северо-Западной Европы, славян Северо-Восточной Европы, альпийцев центральной Европы, средиземноморцев Южной Европы, северной Африки и ближнего Востока, и индостанцев Индии и Пакистана; (2) аборигены восточной Азии, подразделяющиеся на тибетцев, северных китайцев, классических аборигенов Восточной Азии (корейцы, японцы, монголы) и эскимосов; (3) аборигены Юго-Восточной Азии, подразделенные на южных китайцев, тайцев, бирманцев, малайцев и индонезийцев; (4) американские индейцы, подразделенные на северных, центральных, южных и огнеземельцев; (5) африканцы, подразделенные на аборигенов Восточной Африки, суданцев, аборигенов Западной Африки, банту, бушменов и пигмеев; (6) аборигены островов Тихого океана, подразделенных на меланезийцев, микронезийцев, полинезийцев и негритосов и (7) австралийские аборигены, подразделенные на народы Murrayian Юго-Восточной Австралии и CarpentarianСеверной и Центральной Австралии. Бейкером (Baker, 1974) была предложена близкая таксономия с семью расами, включающая пять главных рас по Блюменбаху и Кой бушменов, представленных готтентотами и бушменами Юго-Западной Африки и пустыни Калахари, а также австралидов, представленных австралийскими аборигенами и меланезийцами.
В 1980-х и 1990-х гг. Ней и Райчаудхури (Nei, Roychoudhury, 1993), а также Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) разработали новый метод классификации человеческих рас на основе ряда генетических полиморфизмов (под полиморфизмом подразумевается, что ген имеет более одной аллели, или альтернативной формы). Методика состоит в том, что данные о частотах аллелей большого числа полиморфных генов, определяющих группы крови, состав белков крови, лимфоцитарных антигенов и иммуноглобулинов в популяциях по всему миру сводятся в таблицы. Табулированые данные затем подвергаются факторному анализу, чтобы найти степень, в которой частоты аллелей связаны и образуют кластеры популяций, генетически сходных друг с другом. Данные Нея и Райчаудхури по 26 популяциям были подвергнуты факторному анализу Дженсеном (Jensen, 1998), выявившим существование в человечестве шести основных групп, которые близко соответствуют расам, выделяемым в классической антропологии. Используя традиционную терминологию это: (1) африканцы районов Африки южнее Сахары (пигмеи, нигерийцы, банту, бушмены); (2) европейцы (саамы, финны, немцы, англичане, итальянцы, иранцы, северные индусы); (3) аборигены Восточной Азии (японцы, китайцы, корейцы, тибетцы, монголы); (4) аборигены Юго-Восточной Азии (южные китайцы, тайцы, филиппинцы, индонезийцы, полинезийцы, микронезийцы); (5) америнды (индейцы Северной и Южной Америки и иннуиты[3]) и (6) австралийские аборигены (аборигены Австралии и Новый Гвинеи).
Такая же методика была использована Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) для анализа большего набора данных по 120 аллелям в 42 популяциях. По этим данным вычислялись генетические различия каждой популяции со всеми остальными. По результатам вычислений было построено дерево генетических связей, группирующее популяции в то, что они назвали «кластерами». Исследователи выявили десять основных кластеров. Это: (1) бушмены и пигмеи; (2) африканцы районов Африки южнее Сахары; (3) аборигены Южной Азии и Северной Африки; (4) европейцы; (5) аборигены Восточной Азии; (6) арктические народы; (7) американские индейцы; (8) аборигены Юго-Восточной Азии; (9) аборигены островов Тихого океана; и (10) аборигены Австралии и Новой Гвинеи. Очевидно, что эта классификация близко соответствует расовым таксономиям классической антропологии, основанным на видимых особенностях окраски кожи, волос, глаз, пропорций тела, длины конечностей ит. п., но по какой-то причине Кавалли-Сфорца, Меноцци и Пиацци предпочли термин «кластеры».
Расовые различия в заболеваемости
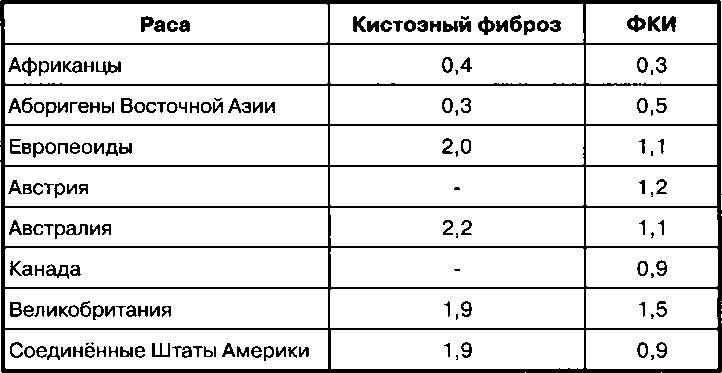
Существуют межрасовые различия по заболеваемости многими болезнями, имеющими генетические корни, в число которых входят кистозный фиброз, фенилкетонурия (ФКИ), гипертония, инсульт, диабет, рак простаты, рак молочной железы, тучность, близорукость и шизофрения. Эти различия возникли вследствие эффекта основателя, дрейфа генов, мутаций и адаптации. Имеется огромный массив исследований на эту тему, изложение результатов которых составило бы целую книгу. В качестве иллюстрации мы приводим частоты встречаемости гена кистозного фиброза и ФКИ у европейцев, африканцев из Африки южнее Сахары и аборигенов Восточной Азии из работы Бодмера и Кавалли-Сфорца (Bodmer, Cavalli-Sforza, 1976). Данные приведены в Таблице 2.1. Числа обозначают частоты встречаемости гена (процент распространенности) в популяции. Как можно видеть, частоты встречаемости гена муковисцидоза среди европейцев в четыре-пять раз выше, чем среди африканцев районов Африки южнее Сахары и аборигенов Восточной Азии, а частоты встречаемости
Таблица 2.1. Частоты встречаемости (проценты) гена кистозного фиброза и ФКИ у европейцев, африканцев районов Африки южнее Сахары и аборигенов Восточной Азии
Существуют ли расы?
С восемнадцатого столетия до середины двадцатого все антропологи, биологи и социологи признавали, что человеческий род состоит из ряда биологически различимых рас. Так, в 1920-х гг. британский антрополог сэр Артур Кейт писал: «Человеческие типы различаются настолько очевидно, что из толпы людей, собранной из австралоидов, негроидов, жителей Восточной Азии и кавказоидов, антрополог смог бы отличить человека одного типа от другого без колебания или ошибки» (Keith, 1922 p.).
Курьёзно, но это кажущееся бесспорным утверждение начало оспариваться с середины двадцатого столетия, когда многие антропологи стали заявлять, что рас не существует. Одним из первых занял эту позицию антрополог Эшли Монтагю (Montagu, 1945а) в своей книге «Самый опасный миф человечества: Заблуждение расы». Название предполагает, что понятие расы — миф, и, следовательно, никаких рас нет. Но из текста книги становится ясным, что для Монтагю расы всё же существуют. Он писал: «В биологии под расой понимают подразделение биологического вида, которое наследует физические особенности, отличающие его от других популяций вида. В генетическом смысле раса может быть определена как популяция, которая отличается по частоте определенных генов от других популяций, с одной или несколькими из которых она способна к обмену генами через барьеры (обычно географические), которые могут разделять их. Если нас спрашивают, существует ли в этом смысле определённое количество рас в человеческом роде, то ответ будет — да» (стр. 6).
Из этого ясно, что раса не «миф» и не «заблуждение». Принимая во внимание то, что Монтагю очевидно признал существование рас, кажется странным, что он дал своей книге такое вводящее в заблуждение название.
Позже, во второй половине двадцатого столетия, многие антропологи и генетики стали утверждать, что рас не существует. В 1962 г. антрополог Ф. Б. Ливингстон (Livingstone, 1962) опубликовал работу: «О несуществовании человеческих рас», в которых он заявил, что «нет никаких рас, есть только переходные формы» (стр. 279). Переходные формы — гибриды между двумя чистыми расами. Переходные формы неизменно появляются в зонах контакта подвергающихся интребридингу рас, производящих расовые-смешанные гибриды. Так, в Латинской Америке проживает большая популяция метисов, имеющих европейских и индейских предков, которая может считаться переходной формой. Аналогично, аборигены островов Тихого океана — переходная форма, возникшая в результате межрасового смешивания аборигенов Юго-Восточной и Восточной Азии. Часто утверждается, что существование промежуточных форм, градиентов признаков, или гибридов лишает правомочности понятие расы. Очевидно, что это не так. Переходные формы и гибриды собак называют полукровками, но существование полукровок не означает, что нет чистых пород.
Но в следующем десятилетии генетики Уолтер Бодмер и Луиджи Кавалли-Сфорца (Bodmer, Cavalli-Sforza, 1976, стр. 698) вынуждены были написать о «существовании многих расовых групп у человека» и что «расы можно назвать подвидами, если мы примем для человека критерий зоологической систематики. Критерием, позволяющим считать две или более групп подвидами, является наличие 75 или более процентов особей, составляющих группы, которые могут быть однозначно классифицированы как принадлежащие к определённой группе». Далее они пишут, что при более широком определении человеческих рас возможно идентифицировать расу при её более чем 75-процентном представительстве в популяции. Следовательно, человеческие расы безусловно существуют. Приблизительно двадцать лет спустя тот же Луиджи Кавалли-Сфорца предпочёл плыть по течению и мы читаем у него о «научной несостоятельности понятия человеческих рас» и что «концепция расы не получила признания» (Cavalli-Sforza, Menozzi, Piazza, 1994, стр. 19). Однако авторы пишут: «Мы можем идентифицировать «кластеры» популяций». Эти кластеры оказываются тем же, что и расы классической антропологии, и далее в своей книге авторы используют классическую расовую терминологию. Например, они пишут, что Африка «населена двумя аборигенными группами, кавказоидами на Севере почти до южных границ Сахары и негроидами в районах Африки южнее Сахары» (стр. 167). Очевидно, они забыли свое предыдущее утверждение о том, что «понятие человеческих рас научно несостоятельно, человеческий род может быть разделен лишь на «кластеры»» (прозрачный эвфемизм для рас). Всего лишь шесть лет спустя тот же Луиджи Кавалли-Сфорца вновь явно передумал, так как заявил, что расы всё-таки существуют, и что раса может быть определена как «группа индивидуумов, которых мы можем распознать как биологически отличных от других» (Cavalli-Sforza, 2000, стр. 25). Возникает впечатление, что он решил отрицать существование рас, но время от времени он забывается, и слово проскальзывает.
К началу двадцать первого столетия отрицание существования рас становится все более и более частым. В 2004 г. Американская Антропологическая Ассоциация объявила на своём Интернет сайте, что «раса не является научно-обоснованной биологической категорией». «Нет никаких биологических рас», утверждает Джефферсон Фиш (Fish, 2002, стр. xii), профессор психологии из Университета Святого Иоанна в Нью-Йорке, но не поясняет, на каких основаниях он делает такое утверждение. Грейвс (Graves, 2002, стр. 2–5), биолог из университета штата Аризона, также утверждает, что «биологические расы не существуют», и пишет, что «термин раса подразумевает существование некоторых существенных основных наследственных особенностей, распространённых в одних группах людей и не представленных в других», и что это не верно для человеческих рас. Вопреки этому утверждению, существует ряд «наследственных особенностей», которые присутствуют в одних расах и отсутствуют в других. Например, гены, определяющие чёрную окраску кожи, присутствуют у африканцев и отсутствуют у европейцев, аборигенов Восточной Азии и американских индейцев, а гены, ответственные за развитие эпикантуса, присутствуют только у аборигенов Восточной Азии, у арктических народов и у некоторых американских индейцев. Более того, понятие расы не обязательно подразумевает, что существуют некоторые аллели (аллели — альтернативные формы генов), которые присутствуют только в одних расах и отсутствуют в других. Достаточно того, что существуют различия в частотах аллели между расами. Для ряда аллелей дело обстоит именно так. Например, аллель серповидно-клеточной анемии встречается намного чаще среди африканцев, чем среди других рас, а аллель кистозного фиброза намного шире распространена среди европейцев (Таблица 2.1).
Грейвс (Graves, 2002, стр. 5) пишет: «Большинство генетиков, эволюционных биологов и антропологов признают, что никаких биологических рас в человеческом виде нет». Коэн (Cohen, 2002, стр. 211) также утверждает: «Почти все антропологи согласны в том, что расы в популярном смысле слова не существуют и никогда не существовали». Эти утверждения не верны. Обзор взглядов американских антропологов, выполненный в 1985 г., показал, что существование рас признавалось 59 процентами биологических и физических антропологов и приблизительно одной третью культурных антропологов (Lieberman и Reynolds, 1996).
Несмотря на отрицание существования рас многими американскими антропологами, реальность понятия расы широко признаётся среди остальной части общества. Медицинские журналы содержат многочисленные статьи на темы расовых различий в заболеваемости и инвалидизации, включая распространенность ВИЧ-инфекции. Издаётся журнал «Этническая принадлежность и здоровье», посвященный расовым различиям в распространенности болезней. В сфере общественных наук есть два журнала, посвященные расовым различиям («Раса и класс» и «Этнические и расовые исследования»). И в других журналах публикуются многочисленные статьи на темы расовых различий в интеллекте, достижениях в образовании, в доходах, социально-экономическом статусе, безработице, предубеждениях, дискриминации, потреблении алкоголя, употреблении табака, наркомании, сексуальном опыте, долголетии, преступности и задержке психического развития. Корпорации способствуют равным возможностям при приеме на работу для представителей разных рас. Служащие предъявляют иски к корпорациям за расовую дискриминацию и часто получают существенную компенсацию по решению судов, у которых нет проблем в понимании того, что такое раса. Многие университеты осуществляют положительную дискриминацию в пользу чернокожих и испано-говорящих соискателей. Судьи выносят решения о неконституционности расово-сегрегированных школ. Граждане многих стран заявляют свою расу в переписных листах, и эти листы анализируются социологами и демографами. В Великобритании есть Комиссия по расовым отношениям, задача которой состоит в том, чтобы способствовать равенству рас и преследовать по суду работодателей за расовую дискриминацию. Ни у людей, ответственных за эту работу, ни у широкой общественности нет трудностей в понимании того, что такое раса, и все они были бы поражены, узнав, что многие американские антропологи утверждают, что рас не существует.
Можно задаться вопросом, почему многие американские антропологи отвергают понятие расы. Ответ был дан двумя польскими антропологами (Kaszycka, Strkalj, 2002, стр. 334). Они пишут:
«Американцы стали чрезвычайно чувствительными к теме расы, и этот термин приобрел крайне щепетильную коннотацию. Многие американские ученые проголосовали за небытие человеческих рас. Кроме того, растущие требования «политической корректности» препятствуют использованию термина и внутри, и вне науки…
Немногие ученые отваживаются заниматься расовыми проблемами, опасаясь получить клеймо расиста только за то, что интересуются этой тематикой».
На причину отрицания понятия расы многими американскими антропологами указывает название книги Монтагю: «Самый опасный миф человечества». Очевидно Монтагю верил, что для людей осознание расы опасно, потому что оно имеет тенденцию питать расовые антагонизмы, которые могут перерасти в конфликт. Для предотвращения этого, было бы лучше понятие расы затушевать. В Европе большинство антропологов принимает правомочность понятия расы. Так, опрос польских антропологов, выполненный в 2001 г., показал что 75 процентов согласны с утверждением: «В пределах вида Homo sapiensимеются биологические расы» (Kaszycka, Strzalko, 2003). Преимущественно в Соединенных Штатах существование рас стало отрицаться многими антропологами и некоторыми биологами и социологами, принесшими свою научную честность в жертву политической корректности.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК