Факты
Что в палеонтологической летописи может служить доказательством эволюции? Таких доказательств несколько типов. Во-первых, наличие широкой эволюционной панорамы: обзор всей последовательности пластов горной породы должен показать, что ранние формы жизни очень просты, а более сложные виды возникли только по прошествии некоторого времени. Кроме того, самые молодые ископаемые, которые мы находим, должны быть ближе всего к ныне существующим биологическим видам.
Мы также должны суметь увидеть случаи эволюционных изменений в ряду поколений, т. е. то, как со временем меняется какое-нибудь растение или животное, превращаясь в нечто иное. Более поздние виды должны обладать чертами, которые делают их похожими на потомков более ранних видов. И, поскольку история жизни включает в себя разделение видов, происходящих от общих предков, мы должны увидеть это разделение в палеонтологической летописи и найти в ней этих предков. Например, анатомы XIX в., основываясь на физиологическом сходстве, выдвинули гипотезу, что млекопитающие произошли от рептилий. Поэтому мы должны найти ископаемые останки рептилий, у которых имеются признаки млекопитающих. Конечно, поскольку палеонтологическая летопись неполна, не стоит надеяться на то, что удастся документировать каждый переход между основными формами жизни. Но по крайней мере часть их мы должны обнаружить.
Когда Дарвин работал над «Происхождением видов», он жаловался на то, что палеонтологическая летопись крайне скудна. В те времена нам не хватало переходных форм ископаемых, недостающих звеньев между основными группами живых организмов, доказывающих существование эволюции. Некоторые группы, такие как киты, возникли в палеонтологической летописи из ниоткуда, и предки их были неизвестны. Но какие-то палеонтологические доказательства эволюции у Дарвина все-таки были. В их число входило наблюдение, согласно которому древние растения и животные разительно отличались от современных видов, и облик их приближался к современным по мере перехода к более поверхностно залегающим, т. е. более поздним пластам горной породы. Дарвин также отметил, что между ископаемыми в соседних пластах больше сходства, чем у тех, которые разделены множеством пластов, и это позволяет предположить, что в прошлом имел место постепенный и длительный процесс дивергенции. Более того, в любом месте ископаемые, найденные в поздних пластах, обнаруживали больше сходства именно с местными современными видами, а не с современными обитателями других краев. Например, ископаемых сумчатых в изобилии находили только в Австралии, а именно там в большинстве своем и обитают современные сумчатые. Это позволяло предположить, что современные виды произошли от ископаемых. (Среди ископаемых сумчатых встречаются самые невероятные млекопитающие, которые когда-либо жили на Земле: например, гигантский трехметровый кенгуру с плоской мордой, грозными когтями и единственным пальцем на каждой задней лапе[15].)
Чего Дарвину не хватало, так это достаточного количества ископаемых, чтобы предъявить четкое доказательство того, как постепенно менялся каждый биологический вид и какие у видов были общие предки. Но со времен Дарвина палеонтологи успели обнаружить множество ископаемых и подтвердили все прогнозы, которые были перечислены выше. В наши дни мы можем показать длительные процессы изменений в ряду поколений; нам хватает доказательств того, что у разных видов были общие предки и переходные формы (например, были обнаружены те самые недостающие звенья в родословной китов); и мы копнули достаточно глубоко, чтобы добраться до самого зарождения сложных форм жизни.
Общие закономерности
Теперь, когда мы расположили все пласты по порядку и провели их датировку, мы можем прочитать палеонтологическую летопись снизу доверху. На рис. 3 показана упрощенная временная шкала истории жизни. На ней отражены главные биологические и геологические события, которые произошли за последние 3,5 млрд лет{6}. Эта шкала показывает четкую эволюционную картину перехода от простых форм к более сложным. Хотя на рисунке указаны моменты возникновения разных групп, например рептилий и млекопитающих, это не означает, что современные формы появились в палеонтологической летописи внезапно, как бы из ниоткуда. На самом деле мы видим, как начиная с ранних форм происходит постепенная эволюция в большинстве групп (скажем, птицы и млекопитающие миллионы лет назад произошли от рептилиеподобных предков). Существование постепенных переходов между основными группами живых существ, о которых я расскажу ниже, означает, что дата возникновения этих групп может быть проставлена довольно-таки условно.
Первые организмы, простые фотосинтетические бактерии, появляются в осадочных породах, насчитывающих около 3,5 млрд лет, т. е. миллиард лет спустя после того, как сформировалась планета Земля. Эти одиночные клетки были единственными обитателями Земли на протяжении следующих двух миллиардов лет, по прошествии которых возникают первые простые эукариоты – организмы, у которых были настоящие клетки с ядром и хромосомами. Затем, примерно 600 млн лет спустя, появляется целый спектр относительно простых, но многоклеточных организмов, в том числе черви, медузы и губки. Эти группы в течение последующих нескольких миллионов лет эволюционируют, порождая многообразие форм живых существ, и 400 млн лет назад появляются земные растения и тетраподы (четвероногие животные, самыми первыми из которых были кистепёрые рыбы[16]). Конечно, более ранние группы зачастую сохранялись: фотосинтетические бактерии, губки и черви появляются в самом начале палеонтологической летописи, и они до сих пор с нами.
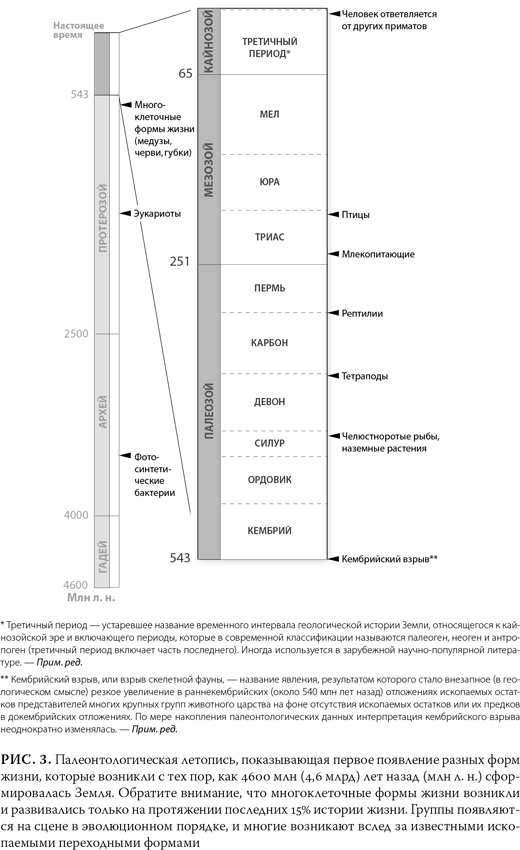
Спустя 50 млн лет мы уже видим первых настоящих амфибий, а по прошествии еще 50 млн лет появляются и рептилии. Первые млекопитающие выходят на сцену около 250 млн лет назад (как и прогнозировалось, они происходят от предков-рептилий), а первые птицы, которые также происходят от рептилий, появляются на 50 млн лет позже млекопитающих. После своего появления первые млекопитающие наряду с насекомыми и земными растениями становятся все более разнообразными, и, по мере того как мы приближаемся к верхним пластам породы, ископаемые все больше напоминают современные виды. Люди на этой сцене совсем новички: наша родословная ответвилась от других приматов всего 7 млн лет назад, и на шкале эволюции это узенькая полоска. Чтобы заострить внимание на этом моменте, проводились самые разные и изобретательные аналогии, но его нелишне будет подчеркнуть еще раз. Если весь ход эволюции представить себе как один год, то самые первые бактерии появятся в конце марта, но первых предков человека мы увидим только в 6 часов вечера 31 декабря[17]. Золотой век античной Греции, около 500 г. до н. э., наступит лишь за 30 секунд до полуночи.
Хотя палеонтологическая летопись растений довольно скудна (ведь у них мало твердых частей, которые превращались бы в окаменелости), но характер эволюции в целом прослеживается сходный. Самые древние растения – это мхи и водоросли, следом появляются папоротники, потом хвойные, затем покрытосеменные (цветковые) растения.
Как показывает палеонтологическая летопись, появление биологических видов разворачивалось далеко не случайным образом. Сложным организмам предшествовало появление простых, предполагаемые предки появились раньше потомков, а самые недавние ископаемые больше всех похожи на современные виды. Многие крупные группы, как выяснилось, связаны между собой переходными формами. И никакой теорией особого сотворения – вообще никакой теорией, помимо эволюционной – эти закономерности не объяснишь.
Окаменелая эволюция и видообразование
Чтобы показать постепенные эволюционные изменения в ряде поколений, требуется хорошая последовательность осадочных пород, желательно таких, которые сменяли друг друга быстро (так что каждый период представлен толстым пластом породы, что позволяет отчетливее проследить перемены). Желательно также, чтобы в последовательности не было пропущенных слоев, поскольку если один из слоев в середине пропущен, то плавный эволюционный переход выглядит как внезапный «скачок».
Идеальным примером служат крошечные морские организмы, скажем, планктон. Таких организмов миллиарды, у многих есть твердый скелет, и они, что очень удобно для исследователя, опускаются после смерти прямо на дно океана, укладываясь в четкой последовательности слой за слоем. Собрать образцы таких организмов, не нарушая порядок слоев, проще простого: нужно лишь пробурить морское дно с помощью длинной трубы, вытянуть наверх цилиндрическую колонку породы (керн) и прочитать (и датировать) его снизу доверху.
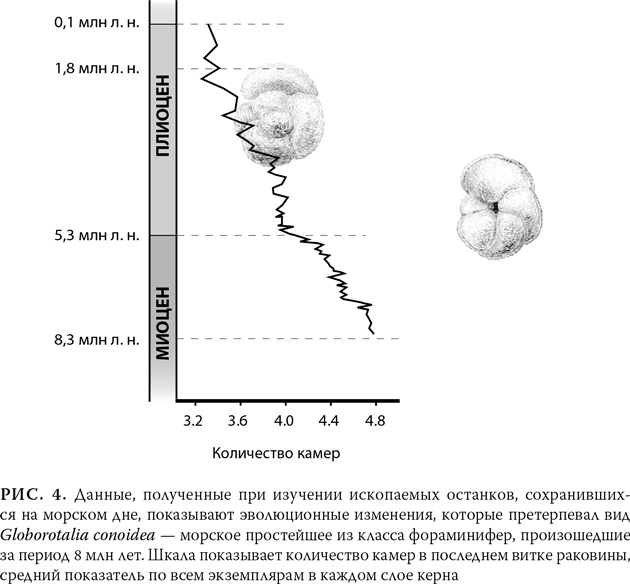
Прослеживая отдельный ископаемый вид с помощью керна, зачастую можно увидеть, как он развивался. На рис. 4 показан пример эволюции крошечного одноклеточного морского простейшего, которое строит спиральную раковину и по мере роста надстраивает новые ее камеры. Образцы взяты из керна длиной 200 м, добытого с морского дна неподалеку от Новой Зеландии, и представляют около 8 млн лет эволюции. Рисунок показывает изменения, которые со временем произошли с одним из отличительных признаков этого простейшего: изменилось количество камер в последнем витке раковины. Здесь с течением времени мы видим довольно плавную и постепенную перемену: в начале у одноклеточного приходится примерно 4,8 камеры на завиток, а к концу – около 3,3 камеры на завиток, т. е. уменьшение составляет около 30 %.
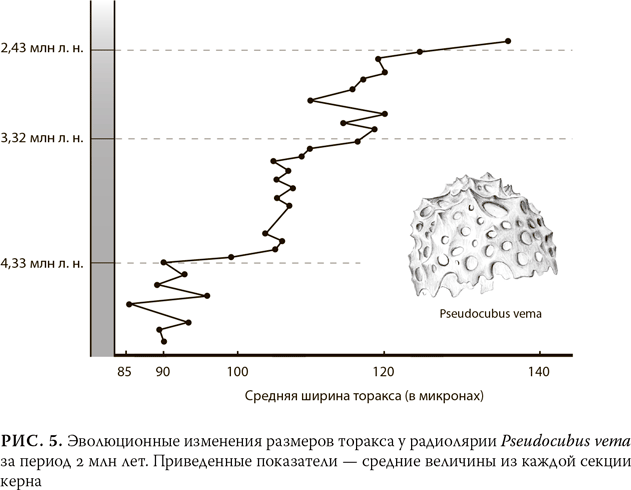
Эволюции, даже постепенной, вовсе не обязательно всегда происходить плавно или равномерно. На рис. 5 показан более неравномерный вариант развития другого морского микроорганизма – радиолярии Pseudocubus vema. В этом случае геологи взяли образцы, располагавшиеся на равном расстоянии друг от друга, из восемнадцатиметрового керна, поднятого с морского дна неподалеку от Антарктиды и соответствующего временно?му промежутку приблизительно 2 млн лет. Признак, который был подвергнут исследованию, – ширина центральной части (так называемый «грудной отдел», торакс) цилиндрического скелета организма, разделенного пережимами на три отдела. Хотя с течением времени размер этого отдела увеличился примерно на 50 %, процесс шел неравномерно. Периоды, когда размер скелета радиолярии практически не менялся, чередовались с периодами более быстрых изменений. Такой тип эволюции часто встречается среди ископаемых останков, и было бы совершенно понятно, если бы подобные перемены были в свое время вызваны факторами окружающей среды, например колебаниями климата или солености. Сами по себе условия окружающей среды меняются нерегулярно и неравномерно, поэтому давление естественного отбора то усиливается, то ослабевает.
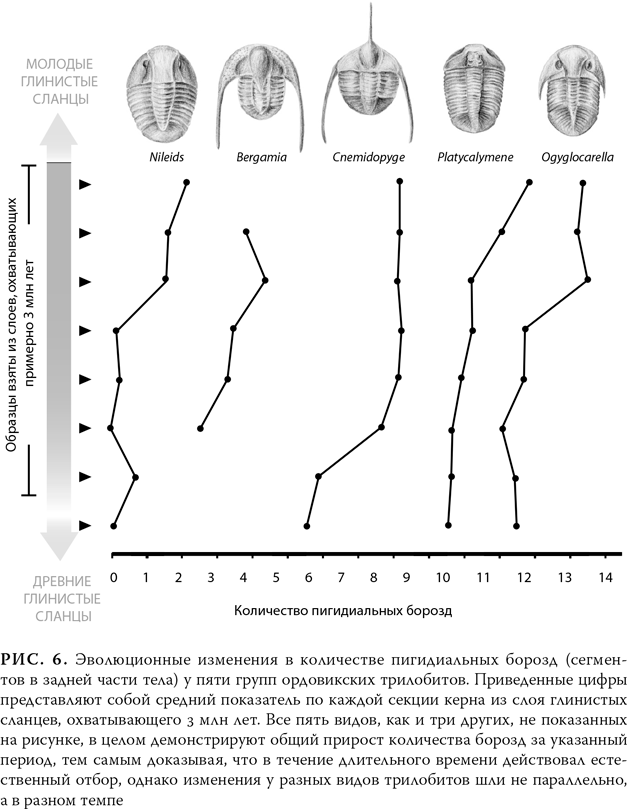
Давайте рассмотрим эволюцию более сложного вида: трилобитов. Трилобиты были членистоногими и принадлежали к той же группе, что насекомые и пауки. Поскольку тело трилобита защищал прочный панцирь, эти ископаемые попадаются в древних породах очень часто (не исключено, что вам удастся купить трилобита в сувенирном магазине при каком-нибудь зоологическом музее). Питер Шелдон, на тот момент работавший в дублинском Тринити-колледже, собирал ископаемых трилобитов в Уэльсе, в слое глинистых сланцев, охватывавшем около 3 млн лет. В этой породе он обнаружил восемь отчетливых линий трилобитов, и с течением времени в каждой из них происходили эволюционные изменения: менялось количество пигидиальных борозд, т. е. сегментов в задней части тела[18]. На рис. 6 показаны изменения у нескольких таких линий. Хотя за весь период, который охватывали образцы, каждый вид в целом демонстрировал прирост количества сегментов, но, если сравнивать разные виды, изменения у каждого из них не только не соотносились с изменениями у другого, но иногда в течение одного и того же периода происходили в противоположных направлениях.
Увы, мы понятия не имеем, какие конкретные воздействия вызвали эволюционные изменения у этих планктонных организмов и трилобитов. Находить свидетельства эволюции в палеонтологической летописи всегда гораздо проще, чем понять, что именно ее вызвало, поскольку, хотя ископаемые и сохранились до наших дней, какова была их среда обитания, неизвестно. Мы можем лишь сказать, что эволюция у них имела место, она была постепенной и у нее менялись как темп, так и направление.
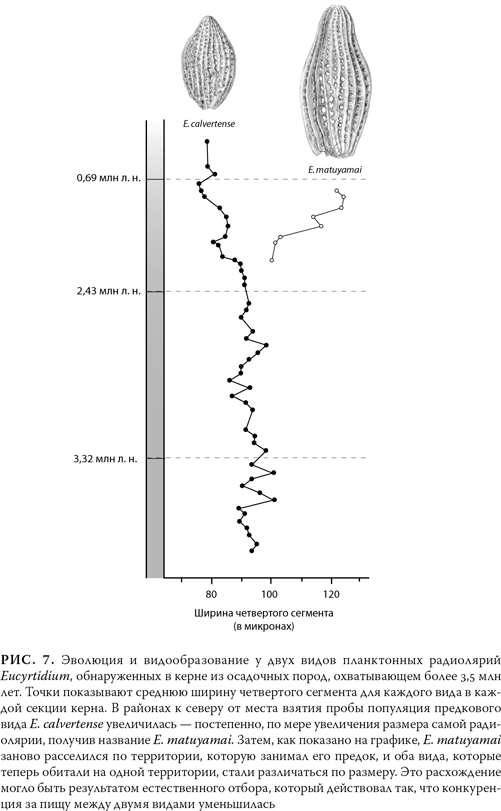
Морской планктон являет собой доказательство как разделения ветвей, так и эволюции в пределах одной ветви. На рис. 7 показано, как предковый вид планктонных радиолярий разветвляется на два других, которые различаются и по форме, и по размеру. Интересно, что новый вид – Eucyrtidium matuyamai сначала сформировался севернее того места, откуда были взяты керны, и лишь потом заселил район, где обитал его предок. Как мы увидим в главе 7, формирование новых видов обычно начинается, когда популяции географически изолированы друг от друга.
Существуют еще сотни доказательств эволюционных изменений в палеонтологической летописи – как постепенных и плавных, так и прерывистых, – причем у таких разных видов, как моллюски, грызуны и приматы. Кроме того, есть примеры видов, которые со временем практически не менялись. (Вспомним: эволюционная теория вовсе не утверждает, что должны изменяться все биологические виды!) Однако, даже если привести еще сотню примеров, мою главную мысль они не изменят: палеонтологическая летопись не дает никаких доказательств в пользу креационистского заявления, согласно которому все биологические виды появились одновременно и затем не менялись. Совсем наоборот: разные формы жизни появляются в палеонтологической летописи в эволюционной последовательности, а затем развиваются и разделяются.
Недостающие звенья
Возможно, изменения у морских видов и служат доказательством эволюции, но это не единственный урок, который преподносит палеонтологическая летопись. Что действительно восхищает, в том числе биологов и палеонтологов, так это переходные формы: те ископаемые виды, которые заполняют промежуток между двумя совершенно разными классами живых организмов. Неужели птицы и впрямь произошли от рептилий, сухопутные животные – от рыб, а киты – от сухопутных животных? Если да, то где же подтверждение из числа ископаемых? Даже некоторые креационисты допускают, что с течением времени могут произойти небольшие изменения в размере и форме – этот процесс именуется микроэволюцией, – однако они отвергают идею, что от одного вида растения или животного может произойти совершенной иной вид (макроэволюция). Сторонники идеи разумного замысла утверждают, что такая значительная разница требует непосредственного вмешательства творца{7}. Хотя в «Происхождении видов» Дарвин не смог указать переходные формы, он бы восхитился тем, как его теорию в наши дни подкрепляют достижения современной палеонтологии. Среди них бесчисленные виды, существование которых было предсказано много лет назад, однако обнаружены и извлечены из земли они были лишь в последние несколько десятков лет.
Но что считать палеонтологическим доказательством для крупного эволюционного события? Согласно эволюционной теории, для каждых двух видов, как бы они ни отличались, когда-то существовал некий вид, который был их общим предком. Этот вид мы можем назвать недостающим звеном. Как мы уже убедились, шансы отыскать этот единственный предковый вид в палеонтологической летописи почти что равны нулю. Палеонтологическая летопись попросту слишком фрагментарна, чтобы мы могли на это рассчитывать.
Однако мы не должны сдаваться, потому что вполне можем отыскать в палеонтологической летописи другие виды, близких родственников нашего недостающего звена, которые не менее веско подтверждают общее происхождение потомков. Приведу один пример. Во времена Дарвина биологи, опираясь на анатомические доказательства, в частности на сходство строения черепа и сердца, заключили, что птицы – близкие родичи рептилий. Тогдашние биологи предположили, что у птиц и рептилий был некий общий предок, который в процессе видообразования породил две ветви потомства: из одной в конечном итоге появились все современные птицы, а из другой – все современные рептилии.
Как мог выглядеть этот общий предок? Интуиция подсказывает нам, что он должен был представлять собой нечто среднее между современной птицей и современной рептилией и обладать чертами, характерными и для тех, и для других. Однако это вовсе не обязательно, как отчетливо показал Дарвин в «Происхождении видов»:
Когда я рассматриваю какие-нибудь два вида, мне трудно преодолеть в себе желание создать в воображении формы, промежуточные непосредственно между этими двумя видами. Но это совершенно неправильная точка зрения; мы должны всегда ожидать формы, промежуточные между каждым данным видом и его общим, но неизвестным предком, а предок, конечно, должен был чем-нибудь отличаться от всех своих модифицированных потомков.
Поскольку рептилии появляются в палеонтологической летописи раньше, чем птицы, мы можем предположить, что общим предком птиц и рептилий была древняя рептилия, и начать искать таковую. В ее облике будут некоторые черты, которые будут свидетельствовать, что она и впрямь была недостающим звеном, давшим две ветви, одна из которых впоследствии положила начало всем современным птицам, а вторая породила динозавров. Подлинно птичьи черты, например крылья и крупная грудина, к которой крепятся летательные мышцы, разовьются лишь позже у той ветви, от которой произойдут птицы. По мере того как эта ветвь развивалась от рептилий к птицам, от нее ответвились несколько видов, которые сочетали в себе черты пернатых и рептилий. Часть этих видов вымерла, а другие продолжили развиваться, пока не превратились в современных птиц. Именно среди этих групп древних видов, родственников видов, находящихся близко к точке ветвления, нам и надлежит искать доказательства того, что у них был общий предок.
Таким образом, чтобы показать, что у двух групп был общий предок, нам не требуется отыскивать и предъявлять ископаемые останки именно того вида, который был их общим предком, или даже вида, находившегося на прямой линии наследования от этого предка к потомкам. Скорее, нам нужно лишь отыскать ископаемые, наделенные чертами, общими для двух групп, а также, что очень важно, нам необходимы хронологические доказательства, показывающие, что эти ископаемые появляются в палеонтологической летописи в нужное время. Переходный вид – это не то же самое, что предковый вид; это просто вид, который сочетает в себе черты организмов, живших до и после него. Учитывая фрагментарность палеонтологической летописи, отыскать эти формы в нужный хронологический период – цель логичная и реалистичная. Например, на переходе от рептилий к птицам переходные формы должны выглядеть как ранние рептилии, но наделенные некоторыми чертами пернатых. Мы наверняка найдем эти переходные ископаемые в том периоде, когда рептилии уже появились, но современные птицы еще не успели возникнуть. Далее переходные формы не обязательно должны находиться на прямой линии наследования от предка к ныне живущему потомку: они могут быть сестринскими эволюционными ветвями, которые вымерли. Как мы увидим, у динозавров, от которых произошли птицы, имелись перья, но некоторые пернатые динозавры продолжали существовать долгое время после того, как появились другие животные, похожие на птиц. Эти поздние виды пернатых динозавров также служат доказательством эволюции, потому что сообщают нам нечто важное о происхождении птиц.
Получается, что датирование и до некоторой степени внешний облик переходных видов отчасти можно предсказать, исходя из эволюционной теории. Часть сравнительно недавних и очень важных прогнозов, которые удалось сделать, касаются нашей собственной группы – позвоночных.
Путь на сушу: от рыб к амфибиям
Одним из крупнейших сбывшихся прогнозов в эволюционной биологии стало открытие переходной формы между рыбами и амфибиями, сделанное в 2004 г. Это ископаемый вид Tiktaalik roseae, который дает нам возможность понять, как позвоночные переселились на сушу. Открытие этого вида – потрясающее доказательство эволюционной теории.
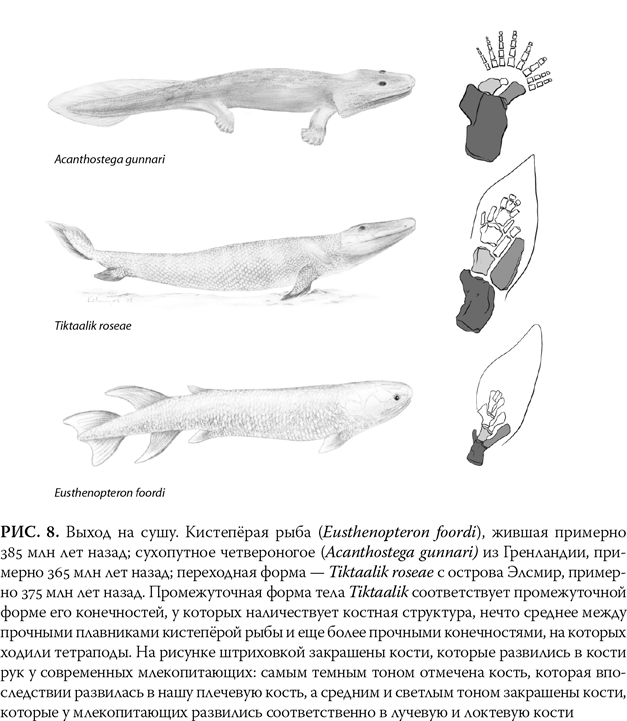
Примерно 390 млн лет назад единственными позвоночными были рыбы. Но 30 млн лет спустя мы уже видим явных животных – тетрапод: четвероногих позвоночных, которые передвигались по суше. Эти ранние четвероногие в некоторых отношениях напоминали современных амфибий: у них были плоские головы и тела, отчетливо выраженная шея, хорошо развитые лапы, а также плечевой и тазовый пояса. Тем не менее у них просматривается и прочная связь с предшественниками-рыбами, особенно с группой, известной как кистепёрые рыбы, названной так из-за крупных костистых плавников, которые позволяли представителям группы ползать по дну мелких озер и ручьев. Ранние тетраподы имели рыбоподобный облик: у них была чешуя, а также схожие с рыбами кости конечностей и головы (рис. 8).
Как эта ранняя рыба развивалась, чтобы получить способность жить на суше? Этим вопросом интересовался, точнее, был одержим мой коллега по Чикагскому университету Нил Шубин. Нил посвятил многие годы изучению того, как из плавников развились конечности, и страстно хотел разобраться в ранних стадиях этой эволюции.
Вот тут на сцену выходит тот самый прогноз. Если 390 млн лет назад существовали кистепёрые рыбы, но не было сухопутных позвоночных, а 360 млн лет назад сухопутные позвоночные уже точно появились, то где искать переходные формы? Где-то между этими этапами. Следуя этой логике, Шубин спрогнозировал, что, если переходные формы существовали, их надлежит искать в слое пород, которому около 375 млн лет. Кроме того, следует искать ископаемые в слое, который образовался из пресноводных, а не из морских отложений, потому что и поздние кистепёрые рыбы, и ранние амфибии обитали в пресной воде.
Изучая карту открытых пресноводных отложений, относящихся к нужному периоду, Шубин и его коллеги методом исключения остановились на палеонтологически неизученном регионе Канадской Арктики – острове Элсмир, который находится в Северном Ледовитом океане, к северу от Канады. И после пяти лет дорогостоящих и бесплодных поисков исследователи наконец нашли то, что искали: группу ископаемых скелетов, покоившихся один на другом в осадочной горной породе на месте древнего пресноводного ручья. Когда Шубин впервые увидел ископаемые останки, показавшиеся из горной породы, он сразу понял, что нашел ту самую переходную форму. В честь местных жителей-эскимосов и благотворителя, который помогал финансировать экспедицию, ископаемое нарекли Tiktaalik roseae (Tiktaalik на эскимосском означает «большая пресноводная рыба», а roseae – зашифрованная отсылка к анонимному дарителю).
Tiktaalik roseae наделен чертами, которые позволяют говорить о том, что он – связующее звено между более ранними кистепёрыми рыбами и более поздними амфибиями (рис. 8). Обладавший жабрами, чешуей и плавниками, он, несомненно, был рыбой, которая жила в воде. Однако у него имеются и черты, присущие амфибиям. Во-первых, голова у него плоская, как у саламандры, а глаза и ноздри расположены скорее в верхней части, чем по бокам черепа. Это позволяет предположить, что тиктаалик обитал на мелководье, мог высовывать голову и, возможно, дышать над поверхностью воды. Плавники у него стали более твердыми и позволяли животному приподняться, чтобы оглядеться по сторонам. Кроме того, у тиктаалика, как и у ранних амфибий, была шея. У рыб шеи нет, их черепа непосредственно соединены с плечами.
Но важнее всего то, что у тиктаалика есть две новые по сравнению с предшественниками черты, которые впоследствии помогли его потомкам переселиться на сушу. Первая – это комплект прочных ребер, которые помогали животному набирать воздух в легкие и перегонять кислород из жабр (тиктаалик мог дышать двумя способами). Вторая – то, что вместо множества маленьких косточек, которые имелись в плавниках у кистепёрой рыбы, у тиктаалика в конечностях костей было меньше и они были прочнее, а по количеству и положению были такими же, как у всех сухопутных существ, возникших позже, в том числе и у человека. В сущности, эти конечности представляли собой наполовину плавник, наполовину лапу.
Ясно, что тиктаалик был хорошо приспособлен для обитания в мелководном пресном водоеме, для того чтобы ползать по дну, приподниматься, выглядывать над поверхностью воды и вдыхать воздух. Учитывая его строение, можно увидеть следующий, решающий шаг эволюции, который, возможно, включал в себя и иные, новые повадки. Некоторые потомки тиктаалика расхрабрились настолько, что выбрались из воды на своих крепких лапах-плавниках, возможно, чтобы перебраться по суше в другой ручей (именно так ведет себя современная необычная тропическая рыба илистый прыгун), спастись от хищников или, быть может, найти пищу среди множества уже возникших в процессе эволюции гигантских насекомых. Если передвижение по суше сулило какие-то преимущества, то естественный отбор мог превратить этих рыб-исследователей в амфибий. Этот первый маленький шаг на сушу оказался гигантским шагом для позвоночных в целом, потому что в конечном итоге он привел к появлению всех сухопутных позвоночных.
Сам тиктаалик не был готов к жизни на суше. Прежде всего у него еще не развились конечности, которые бы позволили ему ходить. Далее, у него все еще были внутренние жабры для дыхания под водой. Поэтому мы можем сделать другой прогноз. Где-то в отложениях пресноводных водоемов, в слое, насчитывающем 380 млн лет, мы найдем одного из первых обитателей суши, у которого были редуцированные жабры, а конечности чуть более прочные, чем у тиктаалика.
Находка ископаемых останков тиктаалика показывает, что наши предки были плоскоголовыми хищными рыбами, которые прятались в неглубоких пресноводных водоемах. Эта находка замечательно связывает рыб с амфибиями. Не менее замечательно то, что открытие этого ископаемого вида не только было ожидаемо, но и также предсказано его точное местонахождение в горной породе определенного типа и возраста.
Лучший способ прочувствовать драматичность эволюции заключается в том, чтобы лично увидеть ископаемые виды или, если есть возможность, подержать их в руках. У моих студентов была такая возможность, когда Нил Шубин принес скелет тиктаалика на занятия, пустил его по рукам и показал, насколько он соответствует описанию подлинной переходной формы. Для студентов этот показ стал наиболее весомым и наглядным доказательством эволюции. Часто ли вам доводилось держать в руках кусочек эволюционного прошлого, тем более такой, который, возможно, был вашим дальним предком?
Подъем в воздух: происхождение птиц
Какая польза в половине крыла? Еще со времен Дарвина этот вопрос задавали, чтобы поставить эволюцию и естественный отбор под сомнение. Биологи говорят нам, что птицы произошли от ранних рептилий, но как могло сухопутное животное развить в себе способность летать? Креационисты утверждают, что естественным отбором этот переход объяснить невозможно, потому что он потребовал бы промежуточных стадий, на которых у животных были бы лишь зачаточные крылья. Они скорее мешали бы живому существу двигаться, вместо того чтобы дать ему эволюционное преимущество.
Однако, если немного подумать, не так уж трудно понять, каковы были промежуточные стадии в эволюции полета, которые имели смысл и приносили пользу тем, кто их проходил. Первая и самая очевидная стадия – это планирование. Планирование возникало в процессе эволюции независимо несколько раз: у плацентарных млекопитающих, у сумчатых и даже у ящериц. Летучие белки очень ловко планируют, расправив складки кожи по бокам: это отличный способ перебраться с дерева на дерево в поисках орехов или спасаясь от хищников. Есть и еще более примечательный «летучий лемур», или кагуан, обитатель Юго-Восточной Азии, у которого есть впечатляющая перепонка, тянущаяся от головы до хвоста. Были случаи, когда удавалось наблюдать полет кагуана на расстояние 138 м, т. е. практически на длину шести теннисных кортов, и при этом в полете животное потеряло лишь 12 м высоты! Так что нетрудно представить следующий этап эволюции: животное, которое махало конечностями, снабженными перепонкой, как у летучего лемура, и благодаря этому совершало настоящий полет, какой мы наблюдаем у летучих мышей. Однако нам уже не нужно ограничиваться только воображением. В нашем распоряжении теперь есть ископаемые виды, которые отчетливо показывают, как появились летающие птицы.
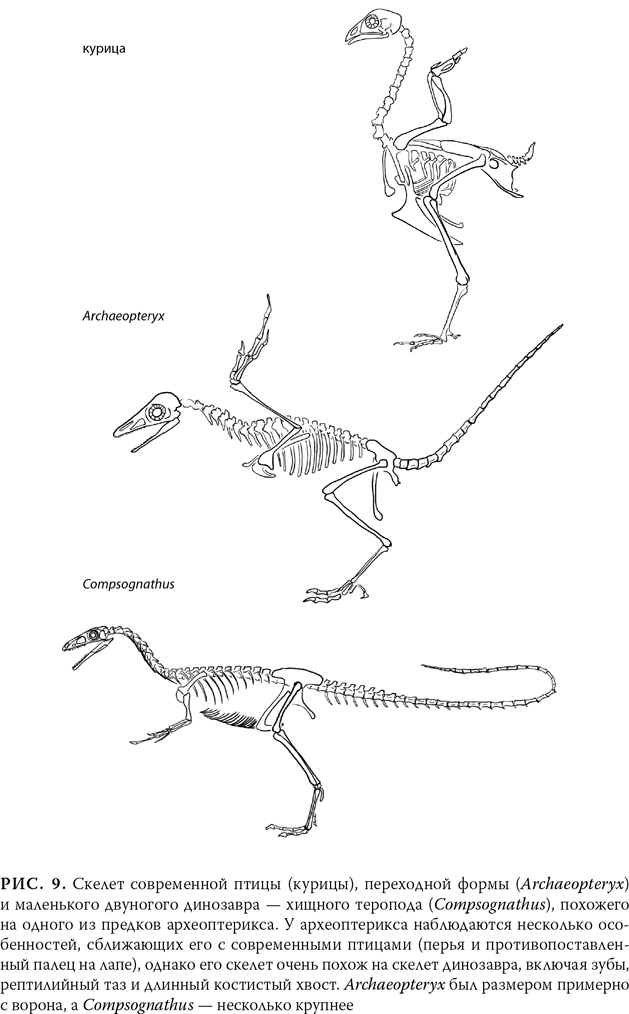
Начиная с XIX в. сходство между скелетами птиц и некоторых динозавров заставило палеонтологов выдвинуть теорию, что у птиц и динозавров был некий общий предок, а именно тероподы – подвижные хищные динозавры, которые передвигались на двух ногах. В палеонтологической летописи примерно 200 млн лет назад встречается немало теропод, но ничего даже отдаленно похожего на птиц. А вот 70 млн лет назад мы уже видим ископаемых птиц, которые весьма напоминают современных. Если эволюция имела место, переходный от рептилий к птицам вид следует искать в промежутке 70–200 млн лет назад.
Там он и обнаруживается. Первое связующее звено между птицами и рептилиями было известно еще самому Дарвину, который, что любопытно, вкратце упомянул о нем лишь в поздних изданиях «Происхождения видов» и то лишь в качестве курьеза. Вероятно, это одна из самых известных переходных форм – Archaeopteryx lithographica, обнаруженный в 1860 г. в Германии, в каменоломне, где добывали известняк. (Название Archaeopteryx обозначает «древнее крыло», а слово lithographica происходит от зольнхофенского известняка[19], который был настолько мелкозернистым, что использовался для изготовления литографических пластин и сохранил даже отпечатки мягких перьев.) Археоптерикс характеризуется как раз таким сочетанием признаков, которые и следовало ожидать от переходной формы. А возраст этого ископаемого вида, около 145 млн лет, помещает его именно в тот отрезок палеонтологической летописи, где и предполагалось его найти.
Археоптерикс был в большей степени рептилией, чем птицей. Его скелет почти идентичен скелету некоторых динозавров-теропод. Что интересно, некоторые биологи, которые недостаточно внимательно рассмотрели ископаемого археоптерикса, не заметили перья и по ошибке отнесли это животное к тероподам. (На рис. 9 показаны общие черты, свойственные этим двум классам.) Характерные признаки рептилий включают челюсть с зубами, длинный костистый хвост, когти, раздельные пальцы на крыле (у современных птиц эти кости слились, как вы можете заметить, если рассмотрите обглоданное куриное крылышко у себя на тарелке) и шею, прикрепленную к черепу сзади (как у динозавров), а не снизу (как у современных птиц). Птичьих черт у археоптерикса всего две[20]: большие перья и противопоставленный большой палец, который, возможно, использовался, чтобы садиться на ветки. До сих пор непонятно, могло ли это существо летать, хотя у него и было оперение. Однако асимметричные перья (у каждого пера одна сторона была больше другой) позволяют предположить, что летать Archaeopteryx мог. Асимметричные перья, подобно крыльям самолета, создают профиль крыла, необходимый для полета. Однако даже если Archaeopteryx мог летать, все равно в основном он был динозавром. Это один из примеров того, что эволюционисты называют «мозаичными признаками»: нельзя сказать, что Archaeopteryx находится на полпути между птицами и рептилиями – с птицами его роднят лишь несколько черт, но большая часть черт у него от рептилий.
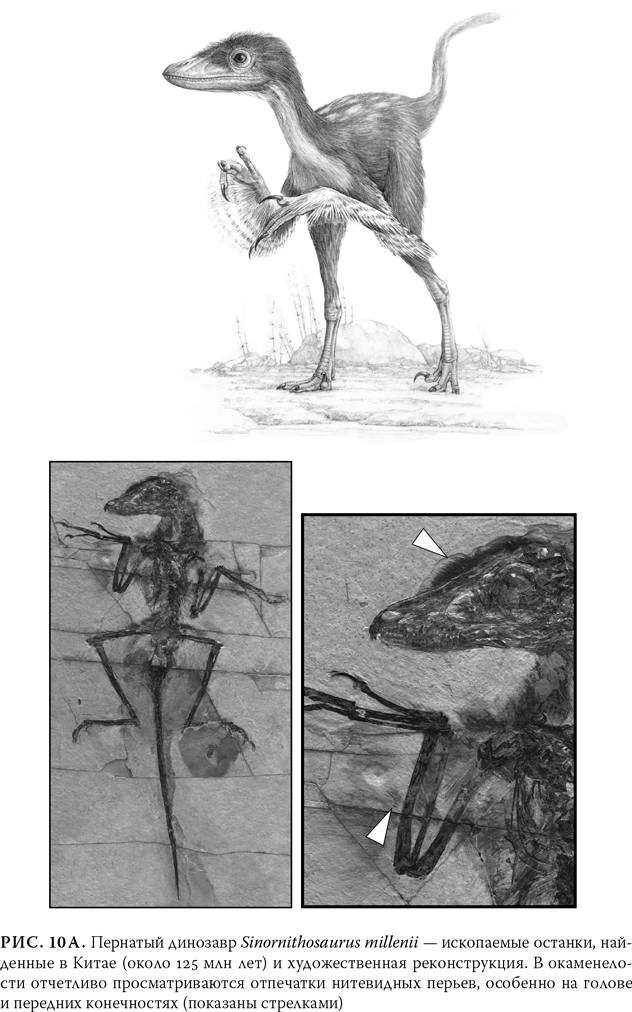
После открытия археоптерикса в течение долгого времени не удавалось обнаружить никаких промежуточных звеньев между рептилиями и птицами, и таким образом между современными птицами и их предками оставалась зияющая брешь. Затем, уже в середине 1990-х гг., брешь начала заполняться благодаря целой лавине поразительных находок, обнаруженных в Китае. Эти окаменелости, обнаруженные в озерных осадочных породах, сохранивших даже отпечатки мягких тканей, являют собой настоящий парад пернатых динозавров-теропод{8}. У некоторых из них все тело покрывали очень мелкие нитевидные структуры – возможно, протоперья. У одного из этих примечательных существ, Sinornithosaurus millenii (Sinornithosaurus означает «китайская птица-ящер»), все тело было покрыто длинными тонкими перьями, такими мелкими, что они вряд ли помогали своему обладателю летать (рис. 10а). А его когти, зубы и костистый хвост отчетливо показывают, что это существо было еще очень далеко от современной птицы{9}. У других динозавров из числа китайских ископаемых на головах и передних конечностях просматриваются перья среднего размера. Однако у других на передних конечностях и хвостах наблюдаются крупные перья, больше напоминающие оперение современных птиц. Самый поразительный из этой компании – «четырехкрылый динозавр» Microraptor gui. В отличие от всех современных птиц это причудливое существо длиной 76 см обладало полностью оперенными передними и задними конечностями (рис. 10б), вытянув которые, по всей вероятности, оно умело планировать{10}.

Динозавры-тероподы, похоже, были не только наделены примитивными птицеподобными чертами, но и некоторыми птичьими повадками. Американский палеонтолог Марк Норелл и группа его коллег описали две ископаемые находки, показывающие поведение динозавров, и если к ископаемым можно применить слово «трогательный», то эти динозавры вели себя именно трогательно. Первая – маленький пернатый динозавр, который спал, спрятав голову под сложенную переднюю конечность, напоминавшую крыло, именно так, как спят современные птицы (рис. 11). Динозавр, получивший название Mei long (что по-китайски означает «крепко спящий дракон»), судя по всему, умер во сне. Вторая – самка хищного динозавра, которая встретила свою смерть, сидя на яйцах, т. е. очевидно, что она высиживала потомство точно так же, как современные птицы.
Все находки ископаемых пернатых динозавров датируются периодом 135–110 млн лет назад, т. е. они жили позже, чем археоптерикс, которому 145 млн лет. Это означает, что они не могли быть его прямыми предками, однако могли быть близкими родственниками. Возможно, пернатые динозавры продолжали существовать и после того, как от одной их эволюционной ветви произошли птицы. Поэтому мы наверняка можем отыскать еще более древних пернатых динозавров, которые были предками археоптерикса. Проблема в том, что перья сохраняются лишь в определенном типе осадочных пород, а именно в мелкозернистых илистых отложениях спокойных водоемов наподобие дна озера или лагуны. А такие условия встречаются очень редко. Однако мы можем выдвинуть еще один эволюционный прогноз, подлежащий проверке: когда-нибудь мы отыщем ископаемые останки пернатых динозавров, которые окажутся старше археоптерикса{11}.
Мы не вполне уверены, что археоптерикс был единственным биологическим видом, от которого произошли все современные птицы. Не похоже на то, что он был недостающим звеном. Но безотносительно к этому археоптерикс представляет собой звено длинной цепочки ископаемых видов (часть которых была открыта неустрашимым Полом Серено), которая отчетливо показывает, как появились современные птицы. По мере того как эти ископаемые виды становятся все моложе, мы видим, как рептилийный хвост у них уменьшается, зубы исчезают, когти срастаются, а также появляется большая грудина, к которой крепятся летательные мышцы.

В сумме эти ископаемые находки показывают, что основная схема скелета и оперение сформировались еще до того, как птицы научились летать. Существовало множество пернатых динозавров, и их оперение совершенно явно родственно оперению современных птиц. Однако если перья появились не как адаптация к полету, то зачем они вообще были нужны? Снова приходится сказать, что ответ на этот вопрос нам неведом. Возможно, у того древнего оперения была своя функция: оно служило для украшения или для демонстрации, чтобы привлечь брачного партнера. Однако более правдоподобным кажется другой вариант: оперение могло служить в качестве теплоизоляции. В отличие от современных рептилий тероподы, вероятно, были отчасти теплокровными, а даже если и не были, то оперение помогало поддерживать температуру тела. Еще загадочнее, откуда и из чего развились перья. Наиболее вероятная версия состоит в том, что перья развились из тех же клеток, из которых у рептилий развивается чешуя, однако с этим предположением согласны далеко не все ученые.
Несмотря на лакуны, мы можем выдвинуть некоторые предположения о том, как с помощью естественного отбора сформировались современные птицы. У ранних плотоядных динозавров удлинились передние конечности с лапами, которые, вероятно, помогали им хватать и держать добычу. Такие хватательные движения должны были способствовать развитию мышц, которые помогали бы передним конечностям быстро вытягиваться и сгибаться обратно, т. е. совершать движение, характерное для настоящего полета, а не планирования. Затем возникло оперение, возможно, для теплоизоляции. С учетом этих эволюционных новообразований можно представить себе по меньшей мере два пути, по которым могло пойти развитие полета. Первый – это так называемый сценарий «с деревьев вниз». Существуют свидетельства того, что некоторые тероподы, по крайней мере частично, обитали на деревьях. Передние конечности, снабженные оперением, помогали бы им планировать с дерева на дерево или с дерева на землю, что позволяло спасаться от хищников, облегчало поиск пищи и смягчало падение.
Второй из возможных и куда более вероятный сценарий называется «с земли вверх». Согласно этому сценарию, полет мог появиться как расширенное продолжение бега и скачков с распростертыми передними конечностями: именно так, скорее всего, двигались пернатые динозавры в погоне за добычей. Кроме того, крылья могли удлиниться, потому что это облегчало бег. Так, азиатский кеклик (или азиатская каменная куропатка), охотничье-промысловая птица, объект исследования Кеннета Диала из Университета штата Монтана, представляет собой ныне здравствующий пример этой стадии. Кеклики почти никогда не летают, а крыльями хлопают, только чтобы легче было взбегать в гору. Хлопанье крыльями не только придает им дополнительное ускорение, но и дополнительное сцепление с землей. Новорожденные птенцы кеклика способны взбегать вверх по склону с уклоном 45 градусов, а взрослые особи взбегают по уклону 105 градусов, т. е. даже не отвесному, а нависающему, и все это только за счет бега и хлопанья крыльями. Преимущество очевидно: умение взбегать на такой крутой склон позволяет кекликам спастись от хищников. Следующим шагом в развитии полета должны были стать короткие подскоки в воздух, какие совершают индейки и перепела, когда чуют опасность.
В обоих описанных сценариях естественный отбор мог начать благоприятствовать особям, которые не просто планировали, совершали броски и прыжки или короткие перелеты, а могли пролететь дальше. Затем могли возникнуть и другие эволюционные новообразования – черты, которые есть у современных птиц, в том числе полые кости, обеспечивающие легкость, и большая грудина.
Детали могут оставаться под вопросом, но существование переходных форм и происхождение птиц от рептилий – это неоспоримый факт. Ископаемые виды, подобные археоптериксу и его более поздним сородичам, демонстрируют сочетание птичьих черт и черт ранних рептилий, и в палеонтологической летописи они приходятся на нужное время. Ученые предсказали, что птицы произошли от динозавров-теропод, и действительно, мы нашли динозавров-теропод с перьями. Выстраивается последовательность во времени – от ранних теропод с телом, покрытым тонким нитевидным пухом, к более поздним, обладателям настоящего оперения, возможно, умевшим планировать. В эволюции птиц мы наблюдаем, как видоизменяются прежние черты (передние конечности с пальцами и тонкими филаментами на коже) и образуются новые (беспалые крылья и перья), как и предсказывает эволюционная теория.
Назад в воду: эволюция китов
Дуэйн Гиш, американский креационист, известен своими яркими и популярными (хотя и крайне ошибочными) лекциями, в которых он обрушивается на эволюцию. Я как-то раз побывал на одной такой лекции, в ходе которой Гиш высмеивал теорию биологов, утверждавшую, что киты произошли от сухопутных животных, родичей коров. Как, вопрошал он, мог случиться подобный переход, если промежуточная форма была бы плохо приспособлена к обитанию и на суше, и в воде и, следовательно, не могла быть создана естественным отбором? (Это напоминает аргумент, связанный с промежуточным половинным крылом, который выдвигают против идеи об эволюции птиц.) В пояснение этого пункта Гиш показал слайд с карикатурой: нарисованным животным, напоминающим русалку, у которой верхняя часть тела была от пятнистой коровы, а нижняя – рыбья. Явно озадаченное своей эволюционной участью, это очевидно неприспособленное животное стояло у самой кромки моря, а над головой у него красовался большой вопросительный знак. Карикатура произвела эффект, на который и рассчитывал лектор: слушатели разразились хохотом. «Какие же тупые эти эволюционисты», несомненно, подумали они.
В самом деле, «корова-русалка» – это нелепый пример переходной формы между сухопутными и водными млекопитающими, «мумудивительный провал», как нарек ее Гиш. Но давайте забудем о шутках, каламбурах и риторике и обратимся к самой природе. Получится ли у нас отыскать какое-нибудь млекопитающее, которое обитает и на суше, и в воде, такое существо, которое, как предполагают креационисты, появиться не могло?
Запросто. Отличным кандидатом станет гиппопотам, который хотя и представляет собой близкого сородича сухопутных млекопитающих, но в то же время настолько близок к жизни в воде, насколько это вообще возможно для наземных млекопитающих. (Есть два биологических вида, карликовый гиппопотам и обыкновенный гиппопотам, научное название которого очень ему подходит – Hippopotamus amphibius). Гиппопотамы проводят значительную часть времени, погрузившись в тропическую реку или болото и наблюдая за своей территорией при помощи глаз, носа и высоко расположенных на голове ушей, и все эти органы чувств они умеют плотно закрывать, когда ныряют в воду. Спариваются гиппопотамы в воде, а их детеныши, которые плавать учатся раньше, чем ходить, рождаются под водой, и матери вскармливают их в воде. Поскольку гиппопотамы по преимуществу водные животные, они особым образом адаптированы к тому, чтобы выходить на сушу и пастись там: питаются гиппопотамы в основном в ночное время, и, поскольку они уязвимы для солнечных ожогов, их кожа выделяет особый секрет, маслянистую красную жидкость – гиппосудоровую кислоту. Эта маслянистая жидкость работает солнцезащитным кремом и, возможно, служит антибиотиком. Из-за нее возник миф, будто гиппопотамы потеют кровью. Гиппопотамы явно неплохо приспособлены к среде обитания, и легко увидеть, что если бы они находили достаточно пищи в воде, то в конечном итоге могли бы превратиться в полностью водных существ, напоминающих китов.
Однако нам нет необходимости напрягать воображение и пытаться представить эволюцию китов, сделав экстраполяцию на основе информации о современных биологических видах. Так сложилось, что родословная китов превосходно прослеживается по богатой палеонтологической летописи благодаря их обитанию в воде и прочным костям, которые хорошо сохраняются в ископаемом состоянии. При этом путь эволюционного развития китов был установлен лишь в последние двадцать лет. Это отличный пример эволюционного преобразования, поскольку в нашем распоряжении имеется череда ископаемых останков, расположенных в хронологическом порядке, возможно, ряд поколений предков и потомков, показывающих, как киты переселились с суши в воду.
Еще с XVII в. было признано, что киты и их сородичи – дельфины и морские свиньи[21] – относятся к млекопитающим. Они теплокровные, живородящие, выкармливают потомство молоком, вокруг дыхал у них растет шерсть. Анализ ДНК кита, а также наличие у некоторых китов таких рудиментарных органов, как тазовые кости и кости задних конечностей, показывают, что предки китов обитали на суше. Киты практически наверняка произошли от парнокопытных, отряда млекопитающих, у которых имеется четное количество пальцев[22] (к ним относятся такие животные, как верблюды и свиньи{12}). Современные биологи считают, что ближайший современный родственник кита – это, как вы уже догадались, гиппопотам, поэтому, возможно, сценарий «из гиппопотамов в киты» не так уж и неправдоподобен.
Однако у китов есть свои уникальные черты, которые обособляют их от сухопутных сородичей. Среди этих черт – отсутствие задних ног, передние конечности в форме ласт, уплощенный хвостовой плавник, дыхало (ноздря в верхней части головы), короткая шея, простые конические зубы (в отличие от сложных больших коренных зубов сухопутных животных), особое строение ушей, позволяющее киту слышать под водой, и мощные выступы в верхней части позвоночника, к которым крепятся сильные мышцы хвоста, участвующие в плавании. Благодаря поразительной серии ископаемых, обнаруженных на Ближнем Востоке[23], мы имеем возможность проследить эволюцию каждой из этих характеристик (за вычетом бескостного хвоста, потому что он не подвергается процессу фоссилизации) от наземных до водных форм.
Шестьдесят миллионов лет назад было множество ископаемых млекопитающих, однако ископаемых китов не было. Существа, напоминающие современных китов, появляются 30 млн лет назад. Таким образом, мы наверняка должны найти переходные формы в пределах этого интервала. Прогноз снова сбывается, и они обнаруживаются именно на этом отрезке. На рис. 12 в хронологическом порядке показаны некоторые ископаемые виды – участники этого эволюционного преобразования, охватывающего период от 52 до 40 млн лет назад.
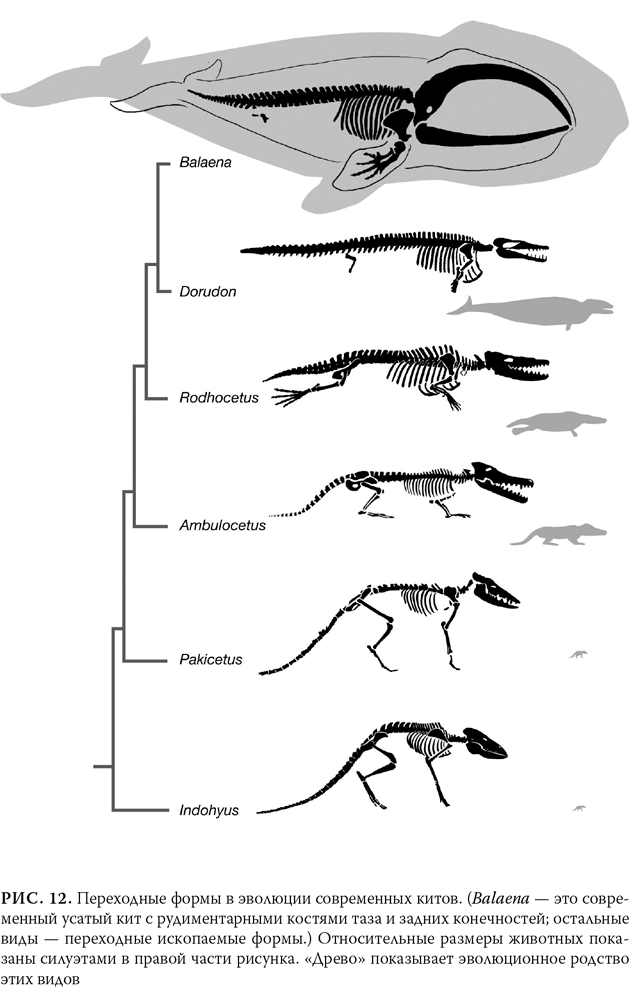
Описывать этот переход в подробностях необходимости нет, поскольку рисунки отчетливо говорят, если не кричат, о том, как наземное животное перебралось в воду. Ряд начинается с недавно обнаруженного ископаемого животного – ближнего родича кита под названием индохиус (Indohyus), которое было размером с енота. Индохиус жил 48 млн лет назад и был, как и предсказывалось, парнокопытным. Его близкое родство с китами несомненно, потому что у него имеются особенности в строении ушей и зубов, которыми наделены только современные киты и их водные предки. Хотя индохиус появляется несколько позже, чем большей частью водные предки китов, он, весьма вероятно, по облику очень близок к тому, как выглядел предок кита. И индохиус был по меньшей мере частично водным обитателем. Нам это известно, поскольку кости индохиуса плотнее и тяжелее, чем кости полностью сухопутных млекопитающих, и это помогало животному держаться в воде не всплывая. Кроме того, изотопы, извлеченные из зубов индохиуса, показывают, что он потреблял много кислорода из воды. Возможно, индохиус плавал в мелководных ручьях или озерах в поисках растительной пищи или укрываясь от врагов, примерно так же, как похожее на него современное животное – африканский оленек{13}. Это временное пребывание в воде, возможно, и привело предка китов на путь к превращению в полностью водное животное.
Индохиус не был предком китов, но практически наверняка был близким родственником этого предка. Однако если мы удалимся еще на 4 млн лет в прошлое, к отметке 52 млн лет назад, то увидим весьма вероятного кандидата в предки кита. Это ископаемый череп животного размером с волка, называемого Pakicetus, который несколько ближе к киту, чем индохиус: у него более простые зубы и уши наподобие китовых. Тем не менее Pakicetus мало напоминал современного кита, поэтому если бы вы оказались в том времени и увидели его, то не догадались бы, что это животное или его близкие сородичи положат начало такой значительной эволюционной радиации[24]. Затем с небольшим интервалом идет серия ископаемых, у которых со временем появляется все больше особенностей строения, свойственных водным животным. На отметке 50 млн лет назад находим примечательного Ambulocetus (буквально «ходячий кит») – обладателя удлиненного черепа и редуцированных, но тем не менее крепких конечностей, которые все еще заканчиваются копытами, показывая, кто является предком этого вида. Это животное, по всей вероятности, большую часть жизни проводило на мелководье, а на суше, скорее всего, неуклюже ковыляло, примерно как тюлень. Еще в большей степени водным был родоцетус (Rodhocetus), живший 47 млн лет назад. Его ноздри сдвинулись к задней части головы, и у него был еще более удлиненный череп. На хребте у родоцетуса имелись толстые выступы, к которым крепились хвостовые мышцы, поэтому, судя по всему, животное хорошо плавало, однако по суше передвигалось с трудом: об этом говорят маленькие таз и задние конечности. Ясно, что родоцетус большую часть жизни проводил в море. Наконец, 40 млн лет назад существовали базилозавр (Basilosauros) и дорудон (Dorudon) – явно полностью водные млекопитающие с короткими шеями и дыхалами на макушке. Они вообще не могли находиться на суше, поскольку их таз и задние конечности полностью редуцировались (у пятнадцатиметрового дорудона длина ног составляла лишь 60 см) и не были соединены с остальным скелетом.
Эволюция китов от наземных животных проходила удивительно быстро: большая ее часть заняла всего 10 млн лет. Это лишь чуть дольше, чем временной отрезок, понадобившийся для эволюционного расхождения нашего вида и шимпанзе, произошедших от общего предка, причем это преобразование потребовало куда меньших изменений формы тела. Тем не менее адаптация к жизни в воде не потребовала появления каких-то совершенно новых черт, лишь модификации уже имевшихся.
Но почему некоторые животные вообще переселились с суши в воду? В конце концов, миллионы лет до этого их предки населяли сушу. Мы не знаем в точности, почему именно произошла обратная миграция, но есть несколько гипотез. Один из возможных ответов: исчезновение динозавров и их свирепых морских сородичей, питавшихся рыбой, – мозазавров, ихтиозавров и плезиозавров. Эти животные, не вымри они, не только соперничали бы с млекопитающими за пищу, но и, что весьма вероятно, пожирали самих млекопитающих. Когда соперники-рептилии вымерли, предки китов, возможно, обнаружили незанятую нишу, свободную от хищников и богатую пищей. Море было готово к их переселению, и до всех благ, которые оно сулило, оставался путь лишь в несколько мутаций.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК